














Application of Tau PFF and Cerebral Organoid Co-Culture in Neurodegenerative Disease Research
 Date de publication :2025-08-12
Date de publication :2025-08-12 Nombre de vues :1484
Nombre de vues :1484

Abstract
Tau pre-formed fibrils (PFFs) are widely used to model the prion-like propagation of tau pathology in Alzheimer’s disease (AD) and related tauopathies. Human induced pluripotent stem cell (iPSC)-derived cerebral organoids provide a physiologically relevant three-dimensional (3D) platform that recapitulates key aspects of human cortical development and organization. In this study, we established an in vitro model by integrating tau PFFs with cerebral organoids to investigate tau pathogenesis and evaluate potential therapeutic interventions.
Introduction
AD is a progressive neurodegenerative disorder characterized by amyloid plaques and neurofibrillary tangles, leading to synaptic loss and cognitive decline. Pathological tau, particularly in its fibrillar form, spreads through the brain in a prion-like manner by seeding aggregation of endogenous tau in recipient neurons. This propagation is strongly associated with disease progression in AD and other tauopathies and is commonly modeled using recombinant Tau PFFs. Cerebral organoids derived from human iPSCs recapitulate critical features of early human brain development, including cortical layer organization, diverse neural cell types, and synaptic activity. Their 3D structure and human origin make them an ideal platform for modeling complex neurodegenerative processes. In this study, we established a co-culture model by introducing tau PFFs into iPSC-derived cerebral organoids to recapitulate human tau pathology in vitro. This model induces hallmark pathological features including tau hyperphosphorylation, aggregation, and glial activation, and provides a human-relevant platform for mechanistic studies and therapeutic screening.
Results
● Cerebral Organoids Contain Abundant Mature Neurons and Glial Cells
To establish a physiologically mature organoid model, human iPSC-derived cerebral organoids (Cat. No. CIPO-BWL002K) were generated using a commercial differentiation kit and cultured for over 100 days. These organoids displayed well-organized laminar structures resembling cortical layers II-VI and expressed key neuronal and glial markers, including glutamatergic, GABAergic, and dopaminergic neurons, as well as astrocytes and oligodendrocytes (Figure 1), indicating advanced maturation and cellular heterogeneity.
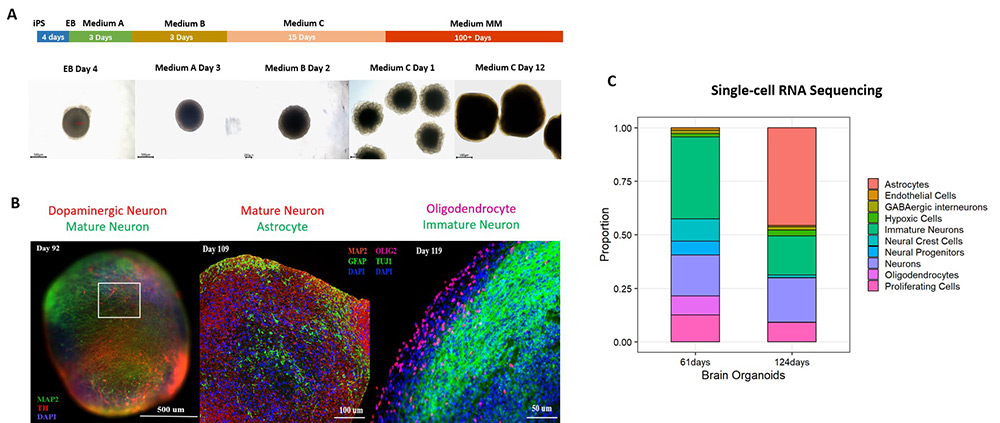
Figure 1. (A) Schematic of the cerebral organoid differentiation process. (B) Immunostaining and (C) single-cell RNA sequencing confirm robust expression of mature neuronal and glial markers in 100+ day cerebral organoids.
● Tau PFF Treatment Induces Cell Apoptosis in Cerebral Organoids
To assess the impact of exogenous Tau seeding, day 100+ organoids were treated with recombinant Tau PFFs (Cat. No. TAU-H5113 / TAU-H5116) at concentrations of 10 μg/mL or 100 μg/mL. The cultures were maintained for 10 days, with medium changes every five days. Apoptotic cells were observed after 5 days of treatment, with noticeable cellular debris surrounding the organoids at 100 μg/mL, indicating a concentration-dependent cytotoxic effect of Tau PFFs (Figure 2).
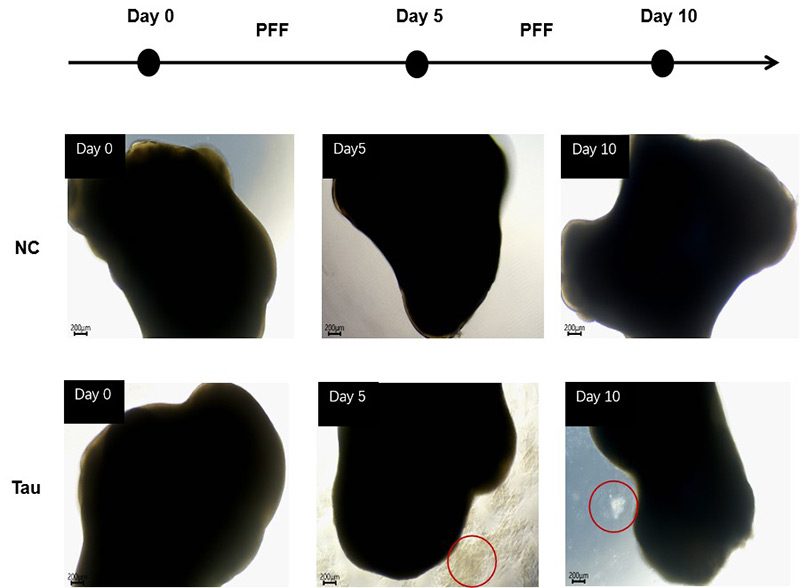
Figure 2. Cell apoptosis was observed after 5 days of Tau PFF treatment.
● Tau PFF Treatment Increases Phosphorylated Tau Levels in a Dose-Dependent Manner
To evaluate the effect of Tau PFFs on Tau pathology, we co-cultured cerebral organoids with recombinant Tau PFFs and assessed phospho-Tau accumulation. Immunostaining revealed significant increases in pTau181 (Figure 3) and AT8 (Figure 4) following treatment. Western blot analysis further confirmed a concentration-dependent increase in pTau217 levels (Figure 5). These results indicate that Tau PFFs induce phosphorylation and seed-dependent propagation of endogenous Tau, replicating key pathological features of AD1-5.
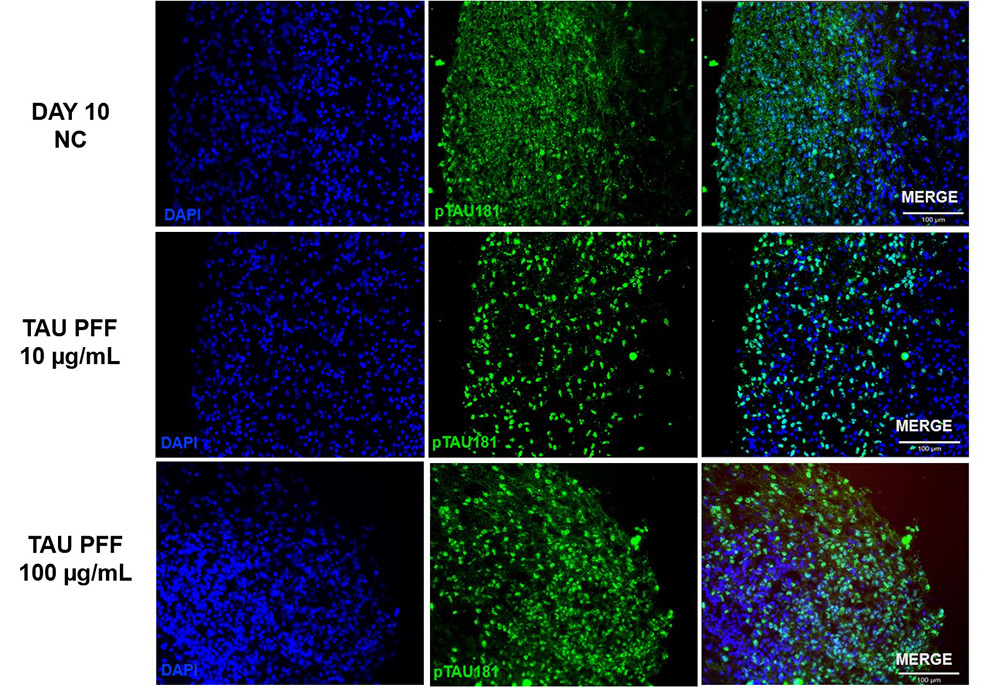
Figure 3. The expression of pTau181 increased following Tau PFF treatment.
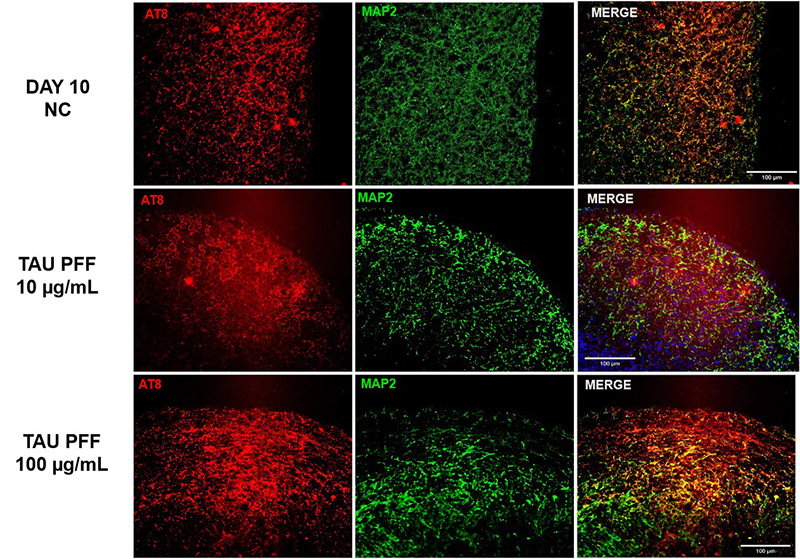
Figure 4. The expression of AT8 increased following Tau PFF treatment.
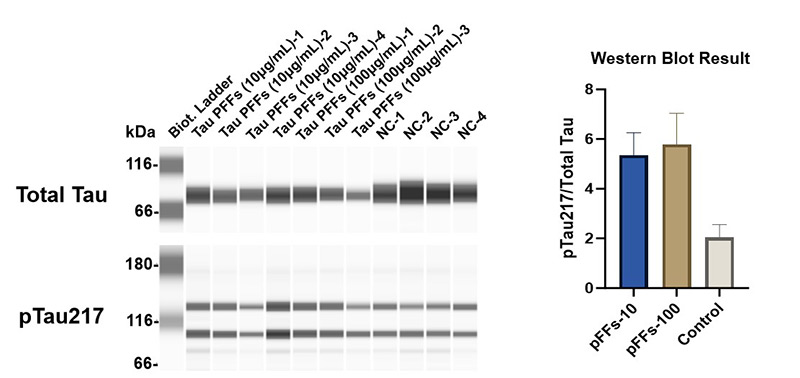
Figure 5. Tau PFFs increase Tau phosphorylation at Thr217 site.
● Tau PFF Treatment Induces Astrocyte Activation in Cerebral Organoids
To evaluate astrocyte activation following Tau PFF treatment, the expression of GFAP and S100β in cerebral organoids was assessed. GFAP expression significantly increased after treatment, particularly at 100 μg/mL (Figure 6), with signals extending from the organoid surface into deeper layers, indicating reactive astrocytosis. S100β, a CNS-specific astrocyte marker, also showed elevated expression and deeper tissue penetration at 100 μg/mL (Figure 7). These findings suggest that high-dose Tau PFFs may induce pronounced astrocyte activation and early neuroinflammatory responses in cerebral organoids5-6.
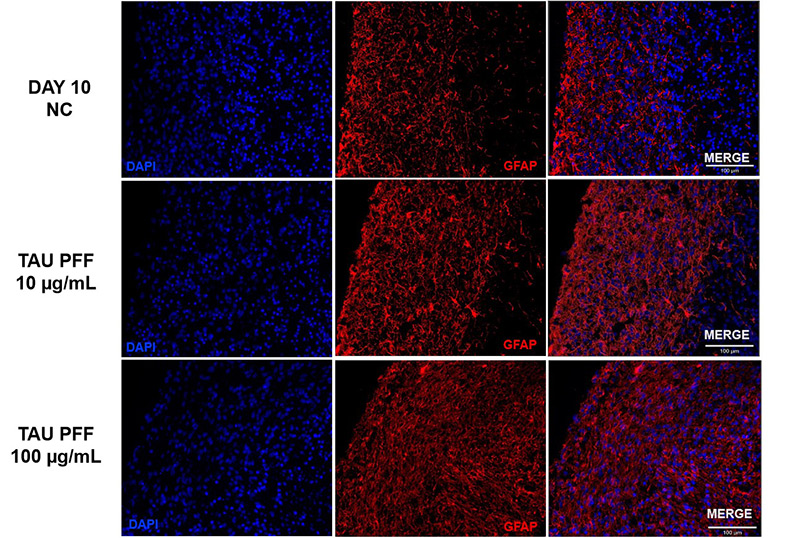
Figure 6. Tau PFF treatment increased GFAP expression, leading to wider distribution and deeper penetration into the brain organoid, indicating enhanced astrocyte activation.
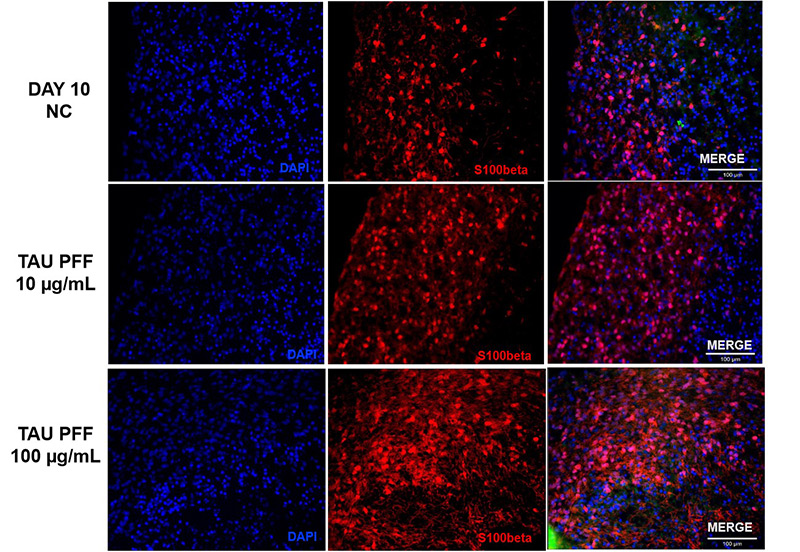
Figure 7. S100β expression also increased after Tau PFF treatment, likely due to astrocyte activation and potential involvement in neuroinflammatory responses.
● Tau PFF Treatment Induces Neuroinflammation and Impairs Antioxidant Defenses in Cerebral Organoids
To investigate the neuroinflammatory and oxidative effects of Tau PFFs, we assessed cytokine expression and antioxidant responses in cerebral organoids. Tau PFF treatment upregulated pro-inflammatory cytokines IL-1β, IL-6, and IL-8, while downregulating SOD2, a key mitochondrial antioxidant enzyme. Elevated IL-1β may activate NF-κB signaling in glial cells, further promoting the release of IL-6 and IL-8. This chronic inflammatory environment may contribute to neuronal injury and enhanced oxidative stress through glia-derived reactive oxygen species (ROS). The downregulation of SOD2 may further impair mitochondrial function by allowing the accumulation of superoxide radicals7-9.
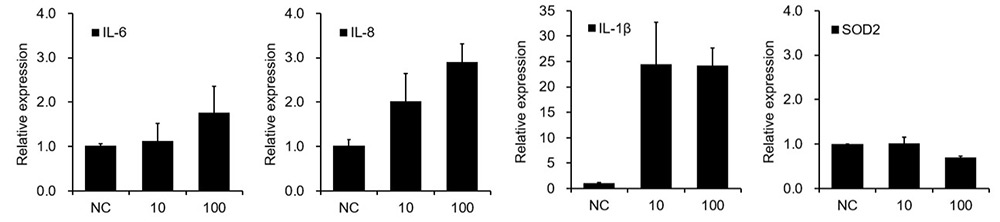
Figure 8. Tau PFF induces neuroinflammation and disrupts antioxidant balance.
Conclusion
This study establishes a robust Tau PFF-cerebral organoid co-culture model that effectively replicates key pathological features of AD and related tauopathies. Exogenous Tau PFFs effectively seed pathology within cerebral organoids, inducing dose-dependent hyperphosphorylation of Tau at disease-relevant sites, including pTau181, pTau217, and AT8. Moreover, Tau PFF exposure leads to pronounced astrocyte activation, as evidenced by elevated expression of GFAP and S100β. The associated pathological changes progressively spread from the organoid surface into deeper cortical regions, reflecting the spatial pattern of astrocyte activation across tissue layers. Activated glia cell may contribute to the pathological cascade by inducing NF-κB-mediated upregulation of pro-inflammatory cytokines such as IL-1β, IL-6, and IL-8, leading to chronic neuroinflammation and increased neuronal vulnerability. The downregulation of the mitochondrial antioxidant enzyme SOD2 further cause ROS accumulation and mitochondrial dysfunction, forming a vicious cycle between inflammation and oxidative stress that exacerbates neuronal damage and apoptosis.
Human-derived organoids bridge the gap between traditional 2D in vitro systems and animal models, providing a pathologically relevant platform for studying Tau aggregation and propagation, neuroinflammation-oxidative stress interactions, and glia-neuron crosstalk. It also enables therapeutic screening for compounds targeting Tau seeding, inflammatory pathways (e.g., IL-1β/NF-κB), or mitochondrial redox imbalance (e.g., SOD2 modulators). Overall, this system offers a powerful tool for mechanistic research and translational studies in neurodegenerative diseases.
Methods and Materials
● Samples and Reagents
Human Tau-441 K18 Pre-formed Fibrils Protein, Tag Free (Cat. No. TAU-H5116)
Human Tau-441 K18 (P301L) Pre-formed Fibrils Protein, Tag Free (Cat. No. TAU-H5113)
Ready-to-use Human iPSC-Derived Mature Cerebral Organoids 100 days+ (Cat. No. CIPO-BWL002K)
Human iPSC-Derived Cerebral Organoid Maturation and Maintenance Kit (Cat. No. RIPO-BWM003)
● PFFs Preparation
Tau PFFs were prepared by incubating recombinant human Tau monomers at 37 °C under constant agitation for 7 days to promote fibrillization. Fibril formation was validated by Thioflavin T (ThT) fluorescence assay and transmission electron microscopy (TEM). The resulting PFFs were aliquoted and stored at −80 °C. Prior to use, PFFs were briefly sonicated to generate shorter fragments suitable for cellular uptake. The sonicated PFFs were then diluted to final working concentrations of 10 μg/mL or 100 μg/mL in cerebral organoid culture medium. Media were exchanged every five days, and treatment was maintained for 10 days.
● Single-cell RNA Sequencing
Single-cell RNA sequencing was outsourced by a third-party provider.
● Immunofluorescence Staining and Simple Western Analysis
Cerebral organoids were fixed with 4% paraformaldehyde, cryoprotected in 30% sucrose, and sectioned using a cryostat. Sections were permeabilized, blocked, and incubated with primary antibodies overnight at 4 °C. After washing, fluorescent secondary antibodies were applied for 2 hours at room temperature. Nuclei were counterstained with DAPI, and slides were mounted using ProLong™ Gold Antifade Mountant. Imaging was performed on a Zeiss Axio Observer Apotome microscope.
For protein quantification, organoids were lysed in RIPA buffer supplemented with protease and phosphatase inhibitors. Protein expression levels of total Tau and phosphorylated Tau (pTau217) were assessed using the Simple Western Jess system (ProteinSimple, Bio-Techne). Sample separation, immunodetection, and chemiluminescent signal acquisition were performed according to the manufacturer’s protocol. GAPDH was used as a loading control. Signal intensities were analyzed and normalized using Compass for Simple Western software (Bio-Techne).
● Real-time quantitative polymerase chain reaction (RT-qPCR)
Total RNA was isolated using the FastPure Cell/Tissue TotalRNA lsolation Kit V2, and template complementary DNA was prepared by reverse transcription using the HiScript lll RT SuperMix for qPCR(+gDNA wiper) (Vazyme). qPCR was performed with the Tag Pro Universal SYBRqPCR Master Mix (Vazyme).
Reference
1. Rawat P, Sehar U, Bisht J, Selman A, Culberson J, Reddy PH. Phosphorylated Tau in Alzheimer's Disease and Other Tauopathies. Int J Mol Sci. 2022;23(21):12841. doi:10.3390/ijms232112841
2. Kaufman SK, Sanders DW, Thomas TL, et al. Tau Prion Strains Dictate Patterns of Cell Pathology, Progression Rate, and Regional Vulnerability In Vivo. Neuron. 2016;92(4):796-812.doi:10.1016/j.neuron.2016.09.055
3. Sanders DW, Kaufman SK, DeVos SL, et al. Distinct tau prion strains propagate in cells and mice and define different tauopathies. Neuron. 2014;82(6):1271-1288. doi:10.1016/j.neuron.2014.04.047
4. Liu F, Huang J, Zhang L, et al. Advances in Cerebral Organoid Systems and their Application in Disease Modeling. Neuroscience. 2019;399:28-38. doi:10.1016/j.neuroscience.2018.12.013
5. Vogel JW, Iturria-Medina Y, Strandberg OT, et al. Spread of pathological tau proteins through communicating neurons in human Alzheimer's disease [published correction appears in Nat Commun. 2021 Aug 5;12(1):4862. doi: 10.1038/s41467-021-25193-3.]. Nat Commun. 2020;11(1):2612. Published 2020 May 26. doi:10.1038/s41467-020-15701-2
6. Michetti F, Clementi ME, Di Liddo R, et al. The S100B Protein: A Multifaceted Pathogenic Factor More Than a Biomarker. Int J Mol Sci. 2023;24(11):9605. Published 2023 May 31. doi:10.3390/ijms24119605
7. Kummer KK, Zeidler M, Kalpachidou T, Kress M. Role of IL-6 in the regulation of neuronal development, survival and function. Cytokine. 2021;144:155582. doi:10.1016/j.cyto.2021.155582
8. Vilotić A, Nacka-Aleksić M, Pirković A, Bojić-Trbojević Ž, Dekanski D, Jovanović Krivokuća M. IL-6 and IL-8: An Overview of Their Roles in Healthy and Pathological Pregnancies. Int J Mol Sci. 2022;23(23):14574. Published 2022 Nov 23. doi:10.3390/ijms232314574
9. Flynn JM, Melov S. SOD2 in mitochondrial dysfunction and neurodegeneration. Free Radic Biol Med. 2013;62:4-12. doi:10.1016/j.freeradbiomed.2013.05.027
 Articles populairesRecommandations associéesActivités populaires
Articles populairesRecommandations associéesActivités populaires
 Why CAR-T Works in Blood Cancers but Struggles in Solid Tumors2026-05-27Nombre de vues :13
Why CAR-T Works in Blood Cancers but Struggles in Solid Tumors2026-05-27Nombre de vues :13 Universal CAR Detection Solutions: Broad Compatibility, Higher Efficiency!2026-05-25Nombre de vues :26
Universal CAR Detection Solutions: Broad Compatibility, Higher Efficiency!2026-05-25Nombre de vues :26 Comprehensive Guide to CAR-T Targets in Solid and Hematologic Malignancies2026-05-21Nombre de vues :157
Comprehensive Guide to CAR-T Targets in Solid and Hematologic Malignancies2026-05-21Nombre de vues :157 Bispecific Antibody Lanscape: Evolution, Breakthroughs, and Prospects2026-05-21Nombre de vues :153
Bispecific Antibody Lanscape: Evolution, Breakthroughs, and Prospects2026-05-21Nombre de vues :153 CD19 as a CAR-T Target: Biology, Limitations, and Alternative Strategies2026-05-20Nombre de vues :128
CD19 as a CAR-T Target: Biology, Limitations, and Alternative Strategies2026-05-20Nombre de vues :128





 Produits associés
Produits associés


